
Hienoa vai karkeaa? Uutta tutkimustietoa raakun poikasten elinympäristövaatimuksista
Jokihelmisimpukan eli raakun pienpoikaset viettävät ensimmäiset elinvuotensa joen pohjasedimenttiin kaivautuneena. Tämä vaihe on raakun elinkierrossa erityisen herkkä ja sen aikainen kuolleisuus on yksi merkittävimpiä syitä raakun uhanalaisuudelle. Alle puolen millimetrin kokoiset poikaset ovat isäntäkalastaan irrottuaan alttiita muun muassa jokipohjan liettymiselle. Joen valuma-alueella tapahtuvan maankäytön seurauksena jokiin päätyvä hienojakoinen maa-aines tukkii hapekkaan pohjasoran, jolloin poikaset tukehtuvat1,2. Kaivautuminen joen pohjaan on kuitenkin raakulle suotuisaa. Se suojaa poikasia saalistajilta (sedimentin pinnalla olevat poikaset maistuvat erityisen hyvin ravuille3) ja estää niitä tempautumasta virran vietäväksi. Pintaan kaivautumista pidetään taas merkkinä poikasten kokemasta stressistä4,5. Sopiva kaivautumisalusta ja kyky kaivautua ovat siis keskeisiä raakun poikasten elossasäilymisen kannalta.

Helmikuussa 2021 julkaistussa tutkimuksessa6 selvitimme, kuinka tarjolla oleva hiekan koko vaikuttaa raakun poikasten kaivautumiskäyttäytymiseen. Sijoitimme raakun poikasia hiekka-alustoille, joiden hiekka edusti yhtä kokoluokkaa: < 120, 120–200, 200–250, 250–500 tai 500–650 mikrometriä. Hiekan koolla oli merkittävä vaikutus poikasten kaivautumiskäyttäytymiseen. Kokeen aikana suurin keskimääräinen osuus kaivautuneita poikasia oli karkeimmassa hiekassa (98 %) ja pienin osuus hienoimmassa hiekassa (52 %). Vaikka raakun poikaset kykenivät kaivautumaan hienoimpaan hiekkaan (< 120 mikrometriä), ne nousivat pintaan muutaman tunnin kuluttua. Poikasilla saattaa olla siis kyky tunnistaa epäsuotuisat olosuhteet – tässä tapauksessa liian hienojakoinen kaivautumisalusta – ja hakeutua niistä pois.
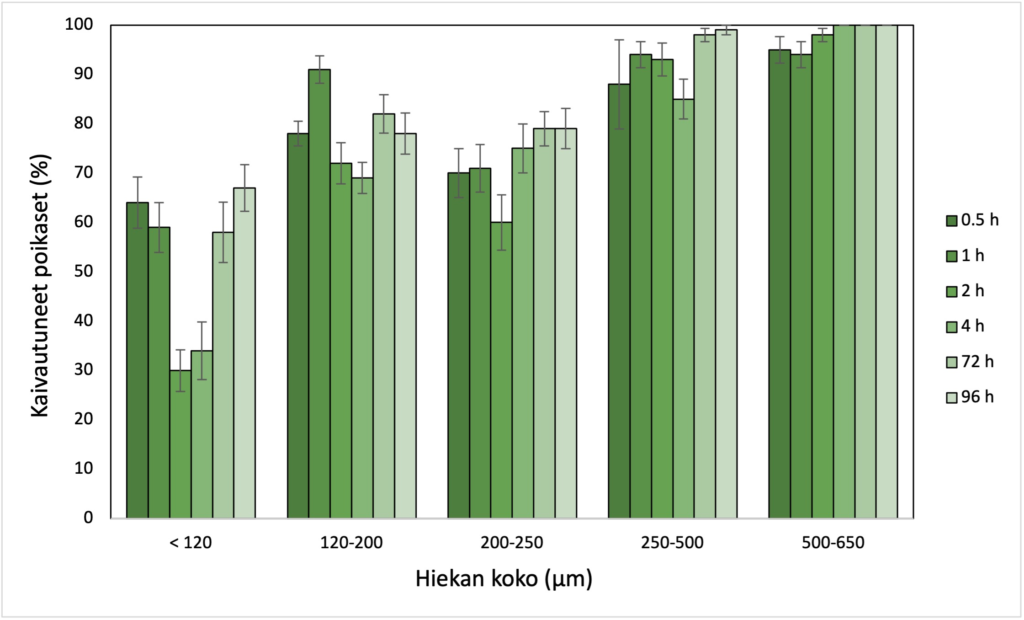
Raakun poikasten kaivautumisaste eri kokoluokan hiekassa. Pylväät edustavat kaivautuneiden poikasten osuutta tietyssä aikapisteessä 30 min – 96 h tarkastelujakson aikana.
Tuloksemme tukevat käsitystä, jonka mukaan jokipohjan koostumus on oleellinen raakun elinympäristön laadun mittari. Koska kaivautuminen on raakun poikasille luontaista ja suotuisaa käyttäytymistä, on tämän kokeen ja siinä käytettyjen hiekan kokoluokkien perusteella 500–650 mikrometrin raekoon hiekka paras kaivautumisalusta raakun poikasille. Tulokset eivät kuitenkaan sulje pois sitä, että karkeampi hiekka tai sora voisi olla poikasille sopiva kaivautumisalusta. Aikaisemmassa tutkimuksessa raakun poikasten on havaittu menestyvän myös 1–2 mm kokoisessa hiekassa7.
Tämän kokeen tulokset ovat uusi askel kohti parempaa ymmärrystä raakun elinympäristövaatimuksista. Raakun uhanalaisimpien populaatioiden keinokasvattaminen laboratoriossa voi olla ainoa keino pelastaa tietyt populaatiot ja säilyttää lajin geneettinen monimuotoisuus, mutta sillä ei voi täysin korvata raakun elinympäristöjen kunnostusta. Ennen keinokasvatettujen raakun poikasten vapauttamista luontoon on tärkeää tunnistaa poikasille sopivimmat elinympäristölaikut purossa. Tuntemalla poikasten vaatimukset ja sietorajat esimerkiksi kaivautumisalustan koostumuksen suhteen, voidaan purokunnostuksia toteuttaa siten, että ne tukevat poikasten elinmahdollisuuksia.
Englanninkielinen artikkelimme raakun poikasten kaivautumiskäyttäytymisestä on luettavissa täällä.
Tämän tutkimuksen mahdollistivat taloudellisella tuellaan Maa- ja metsätalousministeriö, Maj ja Tor Nesslingin Säätiö, Ähtävänjokisäätiö, Suomen Kulttuurirahaston Keski-Suomen Rahasto (Heini Hyvärinen), Suomalainen Konkordia-liitto ry (Heini Hyvärinen), Suomen Luonnonsuojelun Säätiö (Heini Hyvärinen), Raija ja Ossi Tuuliaisen Säätiö ja EU Kolarctic Cross Border Collaboration CBC Programme 2014–2020 (Project SALMUS/KO1017).
Kirjoittaja on väitöskirjatutkija Jyväskylän yliopiston Bio- ja ympäristötieteiden laitoksella.
1 Geist, J. & K. Auerswald. 2007. Physicochemical stream bed characteristics and recruitment of the freshwater pearl mussel (Margaritifera margaritifera). Freshwater Biology 52: 2299–2316.
2 Österling, M. E., B. L. Arvidsson & L. A. Greenberg. 2010. Habitat degradation and the decline of the threatened mussel Margaritifera margaritifera: influence of turbidity and sedimentation on the mussel and its host. Journal of Applied Ecology 47: 759–768.
3 Klocker, C. A. & D. L. Strayer. 2004. Interactions among an invasive crayfish (Orconectes rusticus), a native crayfish (Orconectes limosus), and native bivalves (Sphaeriidae and Unionidae). Northeastern Naturalist Eagle Hill Institute 11: 167–178.
4 Sparks, B. L. & D. L. Strayer. 1998. Effects of low dissolved oxygen on juvenile Elliptio complanata (Bivalvia: Unionidae). Journal of the North American Benthological Society 17: 129–134.
5 Archambault, J. M., W. Gregory Cope & T. J. Kwak. 2013. Burrowing, byssus, and biomarkers: behavioral and physiological indicators of sublethal thermal stress in freshwater mussels (Unionidae). Marine and Freshwater Behaviour and Physiology 46: 229–250.
6 Hyvärinen, H., Saarinen-Valta, M., Mäenpää E. & J. Taskinen. 2021. Effect of substrate particle size on burrowing of the juvenile freshwater pearl mussel Margaritifera margaritifera. Hydrobiologia, 848 (5), 1137-1146. DOI: 10.1007/s10750-021-04522-z
7 Bíly, M., O. Simon, V. Barak & V. Jahelkova. 2020. Occurrence depth of juvenile freshwater pearl mussels (Margaritifera margaritifera) in a river bed tested by experimental mesh tubes. Hydrobiologia. https://doi.org/10.1007/s10750-020- 04298-8.


